|
|
|
Adverts above are not related to this site
AN ACCOUNT OF THE AMPHIBIAN SPECIES FOUND ON POLILLO ISLAND, PHILIPPINES.
Katie Hampson.
Dept Zoology, Oxford University, Oxford OX1 3PS email: khampson@mcmail.com
The following accounts are based upon both anecdotal observations and information gathered from the intensive surveys comparing amphibian assemblages (discussed later). In some cases illustrations of the feet are provided for reference to webbing and tubercle arrangement, which are useful taxonomic features. Any measurements taken are included for reference (Snout Vent length is abbreviated to SV and the mass is in grams). Information on the calls of most species is provided in the sound analysis section. The species accounts provide an introduction and background to the amphibian assemblage comparisons.
Rana vittigerra
This species is frequently seen and heard in rice paddies and amongst coconut plantations. It was not found to breed in streams, therefore probably requires standing water. It was only found at agricultural sites and therefore identified as an agricultural specialist species. Rana vittigerra is found in the Philippines and parts of South East Asia, further work is necessary to clarify the nomenclature and taxonomy of this group.
Bufo marinus
The marine toad, known as the Mindoro frog in the Philippines, is found in rice paddies and around human habitation. Originally a forest species from Mexico, this toad has been introduced to different countries around the world as a form of pest control to prevent crop damage, particularly for protection of sugar cane. In different parts of the world it has spread rapidly and devastated native fauna. On Polillo the marine toad was found in exceptionally large numbers, but never in forest. The toad has been on Polillo for at least 15 years, and is found in the North and South of the island. The reason why the marine toad does not live in forest is not known, but it is not due to limited dispersal capabilities. The marine toad was identified as an agricultural specialist species. There was no evidence that the toad affects existing wildlife in forest on Polillo, any effect on agricultural species cannot be determined without knowledge of the native faunal composition before the toads’ arrival. It is not known to have replaced any native Philippine amphibians (Alcala and Brown 1998), and it is not found in primary forest elsewhere in the Philippines.
Polypedates leucomystax
The common tree frog is often found on banana trees, hence it’s other common name, the banana frog. This species is found most frequently near human habitation and agricultural land, amongst trees and vegetation usually a couple of metres above the ground, but sometimes seen calling in rice paddies perched on the rice stems. A foam nester, it produces large egg masses overhanging shallow water bodies. It was identified as an agricultural specialist species. One adult male was 27g SV 79mm. This species is widespread throughout South East Asia.
Kaloula picta
The fossorial Kaloula picta, (known as the single digit chorus frog) is most active after rainfall. Found by residential areas, rice fields and ponds close to human habitation. This species congregates in large numbers after rain beside water bodies and the chorus of this frog is almost deafening.
Measurements from two frogs are: 8.8g SV 46 mm, 8.5g SV 45mm. Refer to illustrations of feet, but note that K. picta has an inordinate amount of variation in the toe webbing (R. Brown. pers. comm.). Kaloula picta is endemic to the Philippines.
Kaloula sp.
One individual (4g SV 35mm) was found in primary forest amongst leaf litter at 7.30pm on 17/7/99. A second (3g SV 30mm) was found in a pitfall trap in secondary forest on the evening of 23/8/99. Refer to illustration of tubercles on feet. This species is a new record for Polillo. It is rare and possibly restricted to forest based upon the two sightings. This species does not correspond to any of the other Kaloula species that are found on the mainland. The taxonomy of this frog is unclear and requires further investigation.
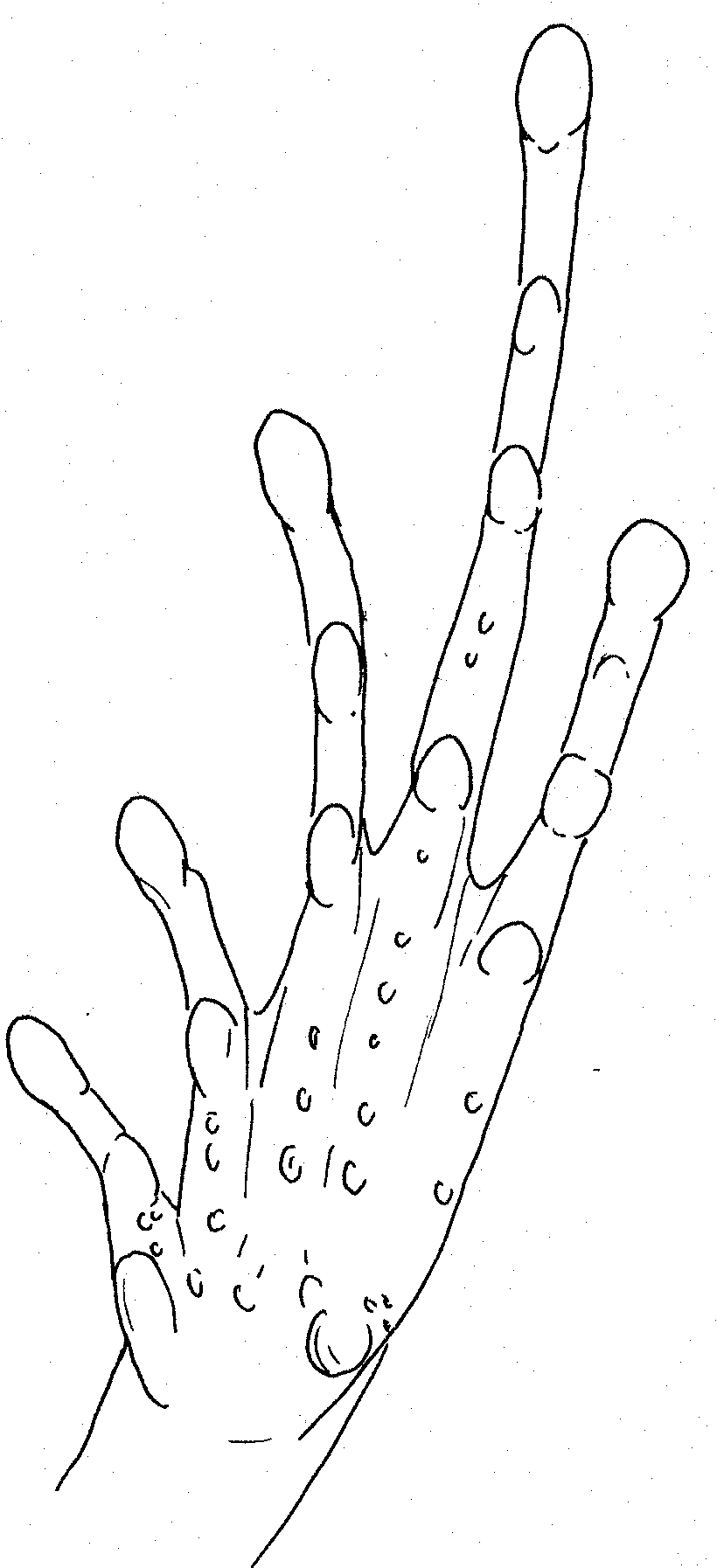
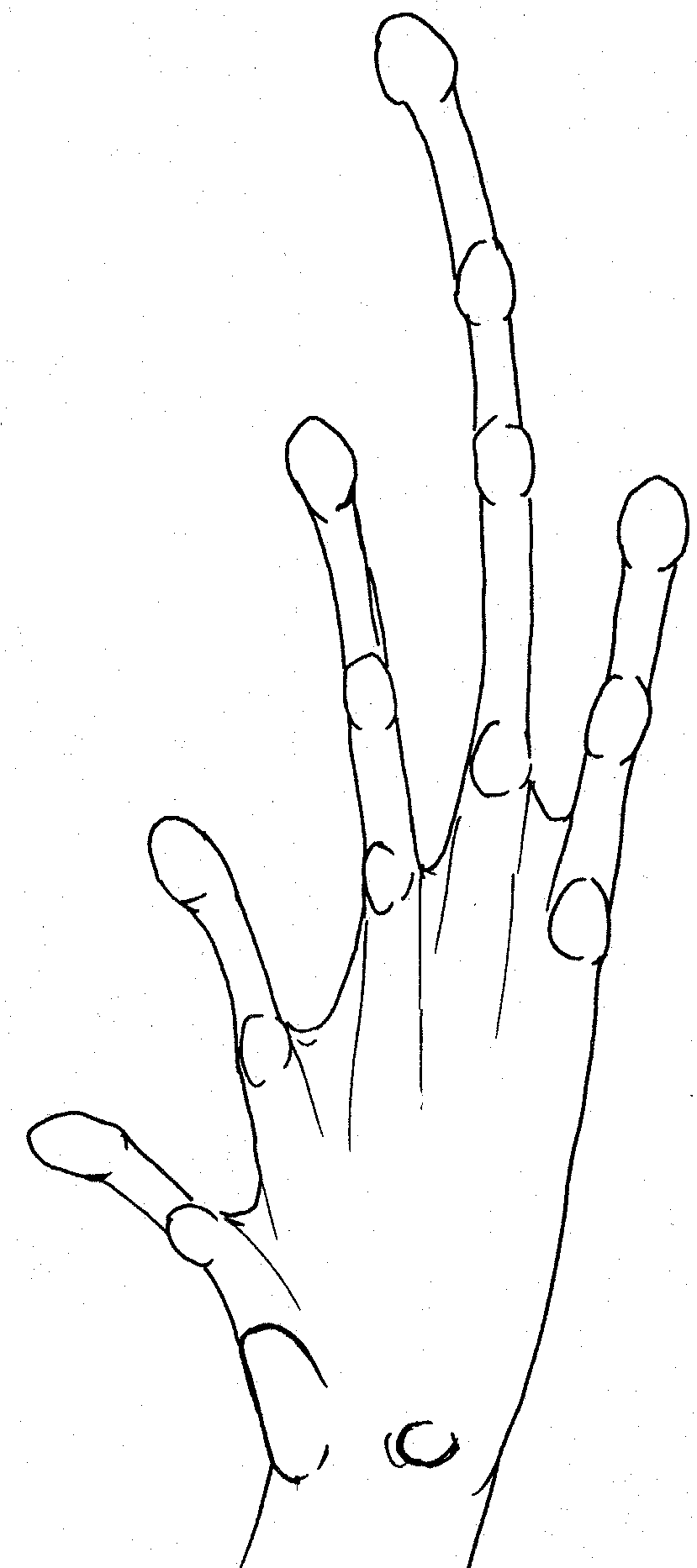
 Right hand
Right hand
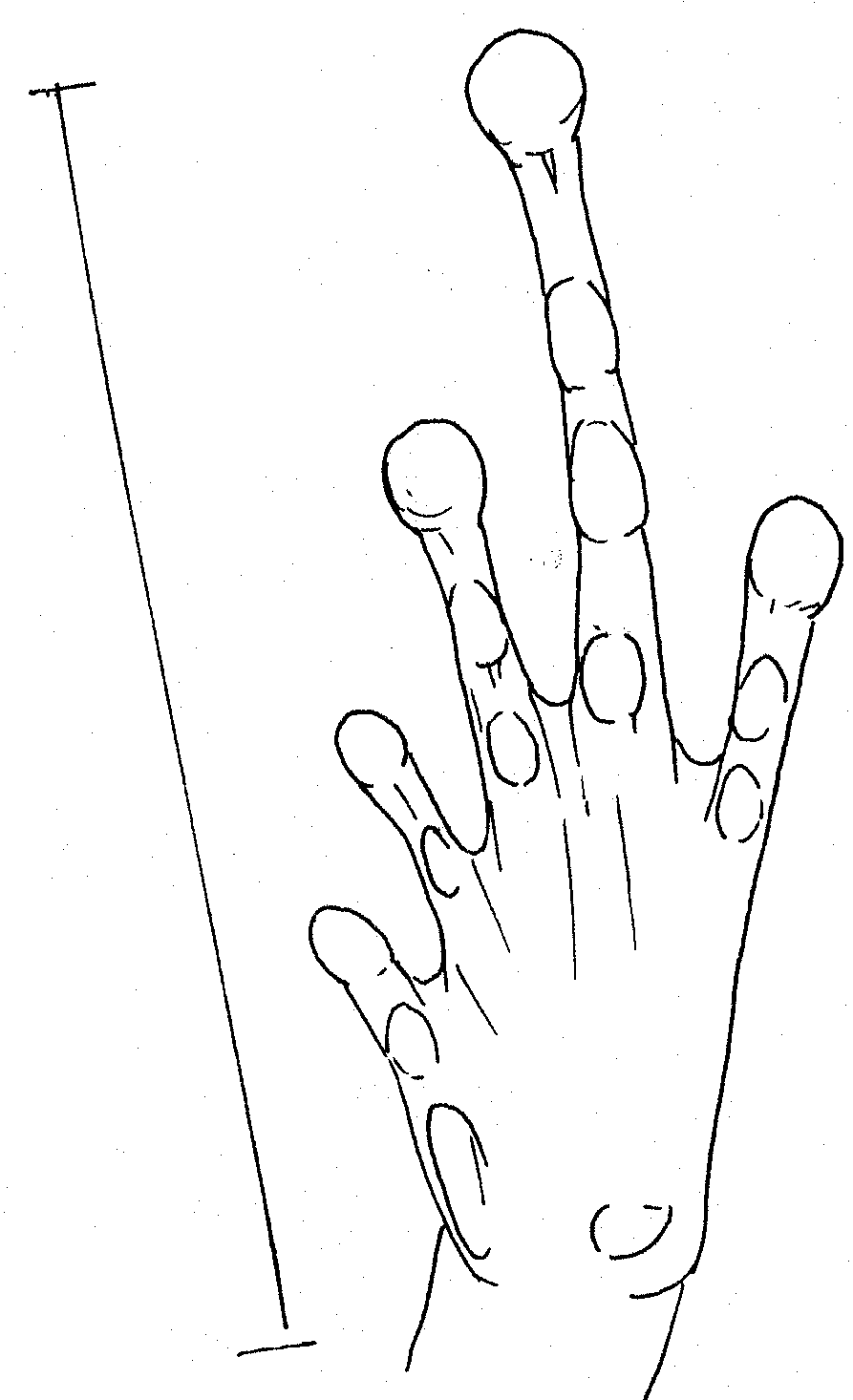
 of Kaloula picta. Scale
13mm Right foot of Kaloula picta. Scale 19mm
of Kaloula picta. Scale
13mm Right foot of Kaloula picta. Scale 19mm
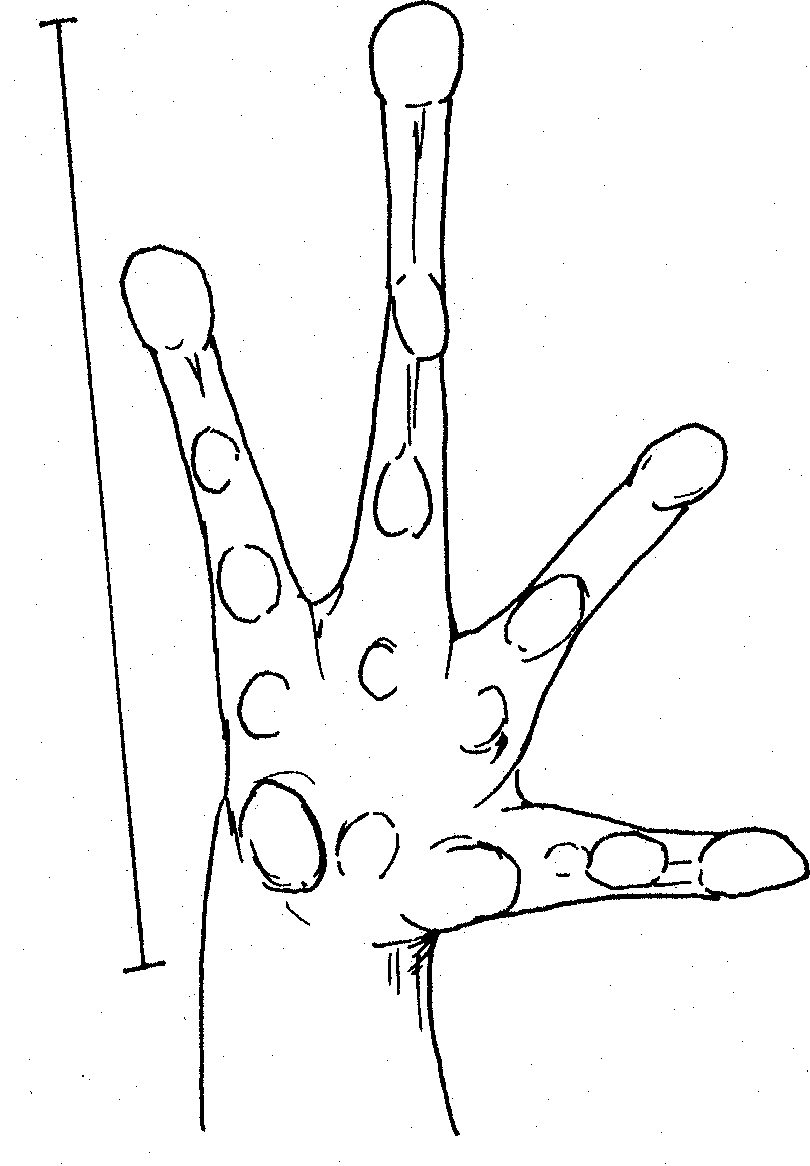
Right hand of Kaloula sp. Scale 10mm.
Very poorly defined tubercles within palm Right foot of Kaloula sp. Scale 15mm
Limnonectes macrocephalus
A large uncommon frog found in low densities. This species is probably more dispersed because of its predatory lifestyle. Its sit and wait tactics are similar to other large ranid species across Asia e.g. Hoplobatrachus tigerinus. It is found in primary forest and also beside large streams in more agricultural habitats. Mean mass 103.4g (n=4, range; 57 to 119.5g), mean SV 121mm (n=4; range 82 to 166mm). The nomenclature of this species has changed, it has been raised to full species level and placed within the Limnonectes genus (R. Brown. pers. comm.). It is endemic to Luzon, Mindoro and the Polillo Islands.
Occidozyga laevis
Is found in most habitats from agriculture to primary forest, excluding rice paddies and can be in large densities especially in very shallow still water. The pattern on this frog is variable. Sometimes either one or two dorsal stripes are present, and sometimes there is a large light patch behind the head. The call is relatively quiet and these frogs are disturbed from calling very easily. This species is widespread throughout the Philippines and parts of South East Asia.
Rana woodworthi
Woodworth’s frog is endemic to a small area of Luzon Island (Laguna and Quezon Provinces) and Polillo. One of the most common frogs on Polillo, particularly abundant in forest and found in agricultural areas in lower numbers.
Rana similis
This frog is commonly seen beside rivers that border forests and rivers within forest. It often sings among rocks or on branches of plants beside the riverbank sometimes a metre or more above the ground. Often heard calling during the daytime in forest. This frog has been designated to full species level (from subspecies Rana signata similis) (R. Brown. pers. comm.). A pair found in amplexus weighed 13.25g SV 59 mm (female), and 4.25g SV 40 mm (male).
Rana luzonensis
This frog is seen on leaves or branches up to 2 metres above the ground and occasionally on the ground. Rana luzonensis was found beside streams in forest, and fast flowing streams among agriculture that are bordered by trees or bamboo. Females often differ in colouration from the males, sometimes brown or beige in colour. Rana luzonensis belongs to the Everetti group and has recently been raised to full species level (R. Brown. pers. comm.). It is endemic to Luzon and the surrounding islands.
A pair in amplexus weighed 6.25g (male), 28.5g (female with approximately 900 eggs). Eggs weighed 3.25g.
Rhacophorus pardalis
This uncommon frog was found in clustered groups. One group was found amongst bushes beside a temporary pool in a coconut plantation, whereas another group was found in forest. A juvenile was also found in forest at site K, and another in a tree at the paddy field site O (see map of frog survey sites later). Mean mass 7.45g (n=2). This species is found in the Philippines, Borneo and Sumatra.
Rhacophorus appendiculatus
One individual was found on the 9/8/99 during the day in primary forest on a tree trunk (3.8g SV 44.5 mm). No others were found on Polillo. However during a visit to Mt Maquiling, near Mud springs, Los Banos (Luzon Island) several individuals were found and heard calling. This species is found in the Philippines, the Malay peninsula, Borneo and Sumatra.
Platymantis dorsalis
This is a very common frog, which calls from the ground and low vegetation. It is heard among coconut groves and forest, in contrast to the mainland where it is found more strictly in forest. It has a non-granular belly, and there are no tiny tubercles over the base of the foot. Refer to illustration of feet. Varying patterns and colouration, sometimes prominent dorsal stripes, either a single central or two dorsal, frequently the dorsal stripe is absent. Platymantis dorsalis is endemic to the Philippines. Mean mass 2.4g (n=16, range; 0.65 to 5.75g), Mean mass of gravid females 5.4g (n=4, 4.95 to 6.2g)
Platymantis corrugatus
 This frog is cryptic, calling
from beneath dead wood and overgrown plants and its distribution is clustered.
This species is found from coconut groves to primary forest. Platymantis
corrugatus is endemic to the Philippines. Tiny tubercles are visible
over base of foot and the belly is granular. See illustrations of feet. Mean
mass 3.25g (n=4, range; 3.0 to 3.85g), Gravid female SV 46 mm 10.5g
This frog is cryptic, calling
from beneath dead wood and overgrown plants and its distribution is clustered.
This species is found from coconut groves to primary forest. Platymantis
corrugatus is endemic to the Philippines. Tiny tubercles are visible
over base of foot and the belly is granular. See illustrations of feet. Mean
mass 3.25g (n=4, range; 3.0 to 3.85g), Gravid female SV 46 mm 10.5g
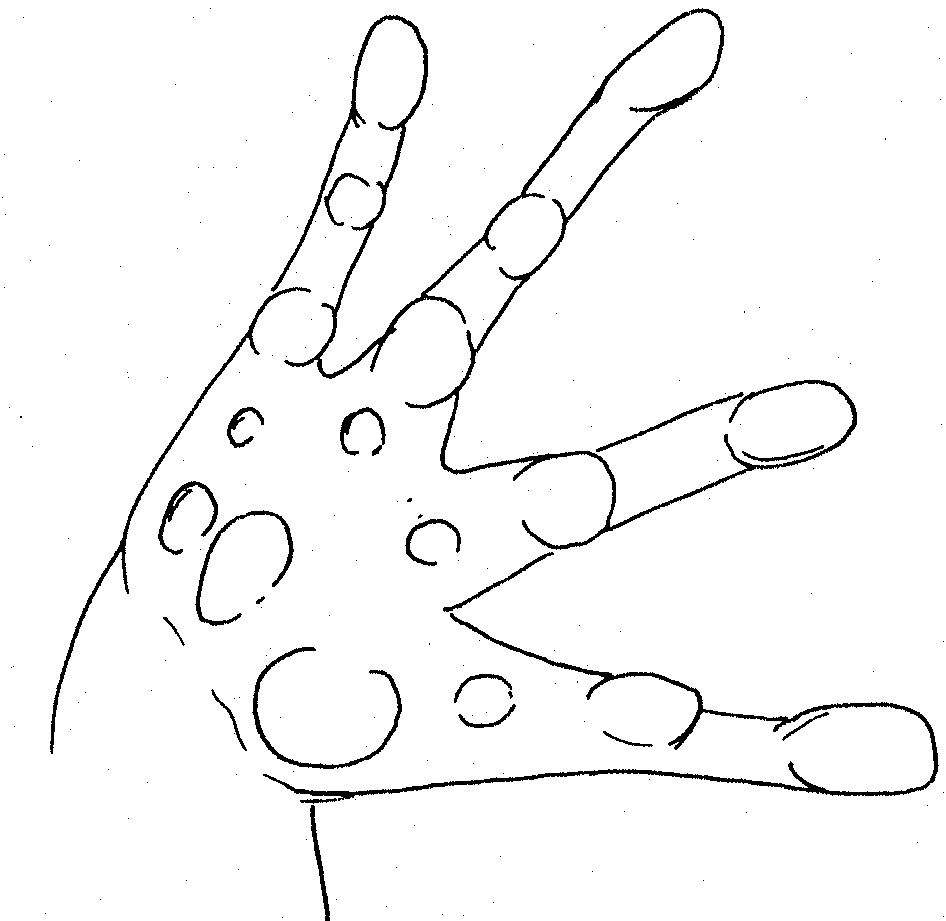
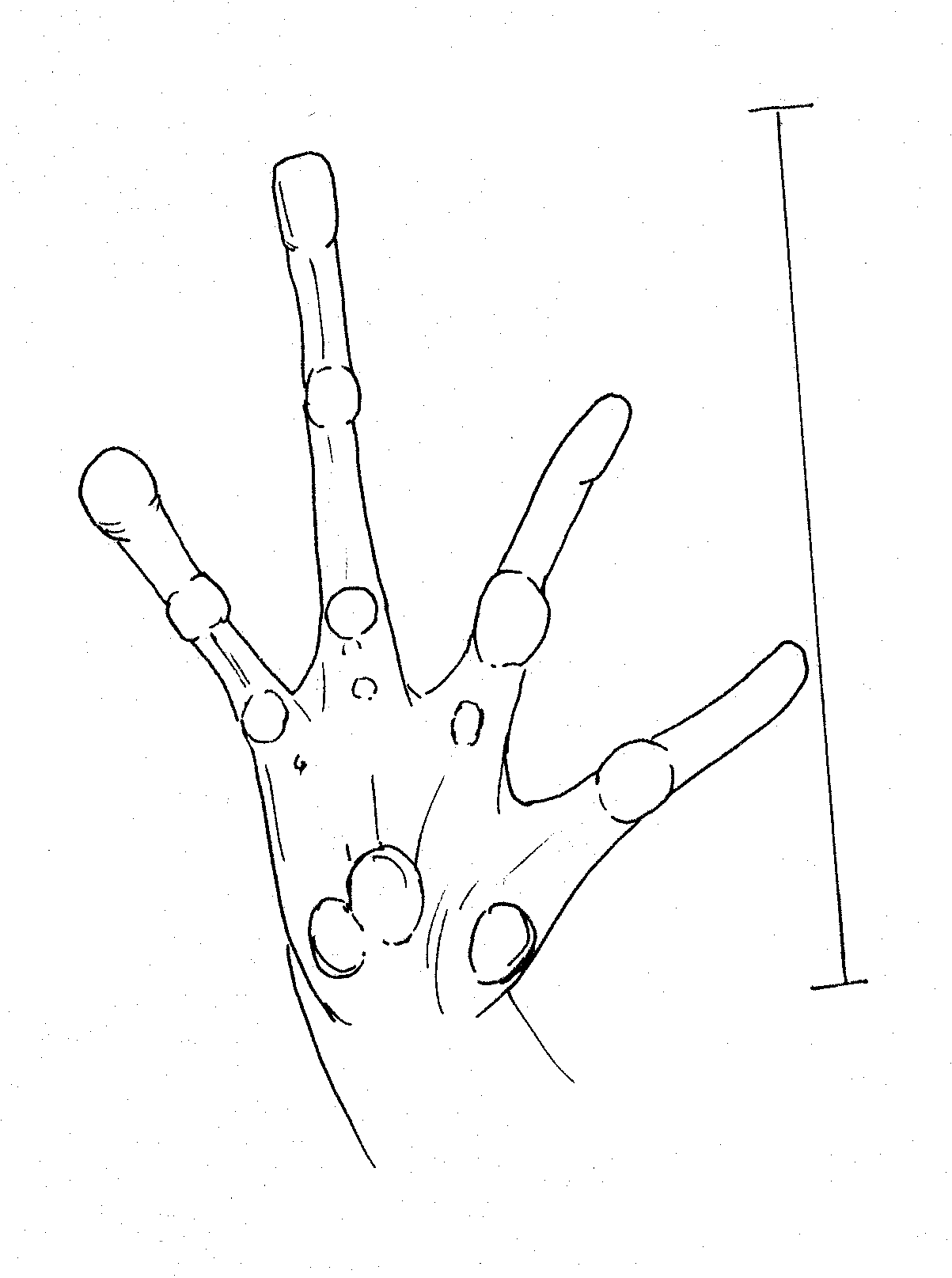
Very prominent tubercles on first two fingers Right foot of female P. corrugatus.
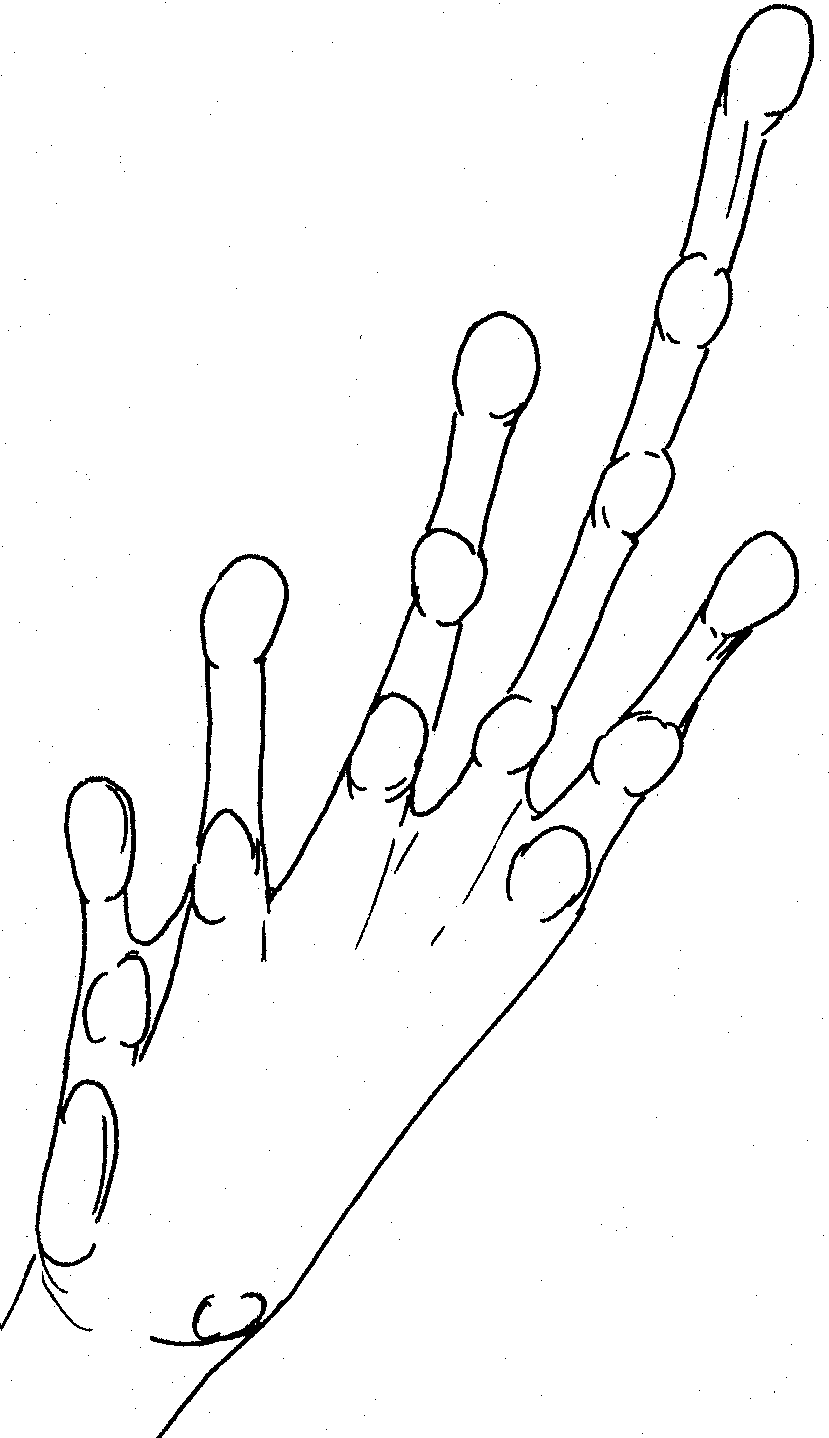 Right
hand of P. dorsalis Right hand of
Right
hand of P. dorsalis Right hand of
P. dorsalis. prominent projecting tubercles
Platymantis sp.
This species has previously been misidentified as the Polillo Forest frog (Platymantis polilloensis). It is now known to be a member of the Platymantis guentheri group. The most similar frog to it on the mainland is Platymantis luzonensis. Platymantis luzonensis has a more warty appearance than Platymantis sp. The calls of the two frogs are very similar (see the sound analysis section). This frog is potentially a different species from P. luzonensis, and possibly endemic to Polillo. Its taxonomy requires further work.
DISTANCE transect data was collected for precise estimations of abundance. These frogs were usually heard calling from at least 2m above the ground, therefore are less likely to be found in transects (see assemblage comparisons) than ground dwelling frogs. According to frequency of calling males these frogs are much less common than both P. dorsalis and P. corrugatus and their habitat is much more restricted to forest. Refer to section on Platymantis sp transects. Mean mass 2.0g (n=3 range 0.8g (juvenile) to 3.05g). Mean female mass 7.09g (n=2). Mean SVL 36.6 mm, (n=7, range 23 to 48)
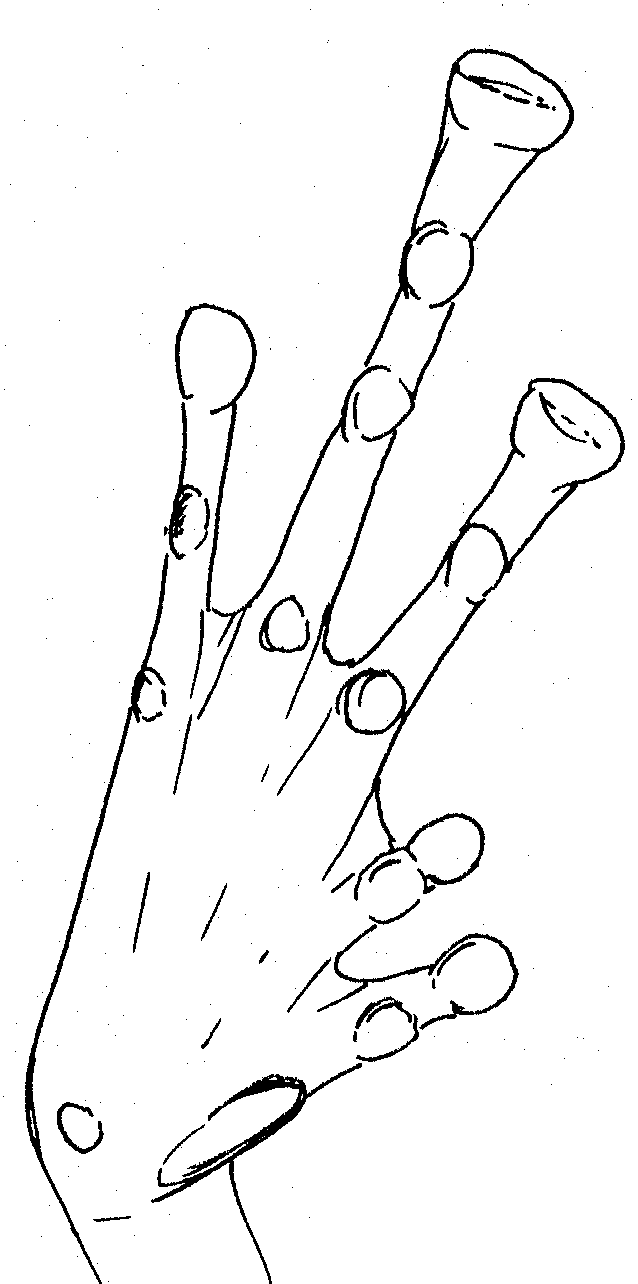
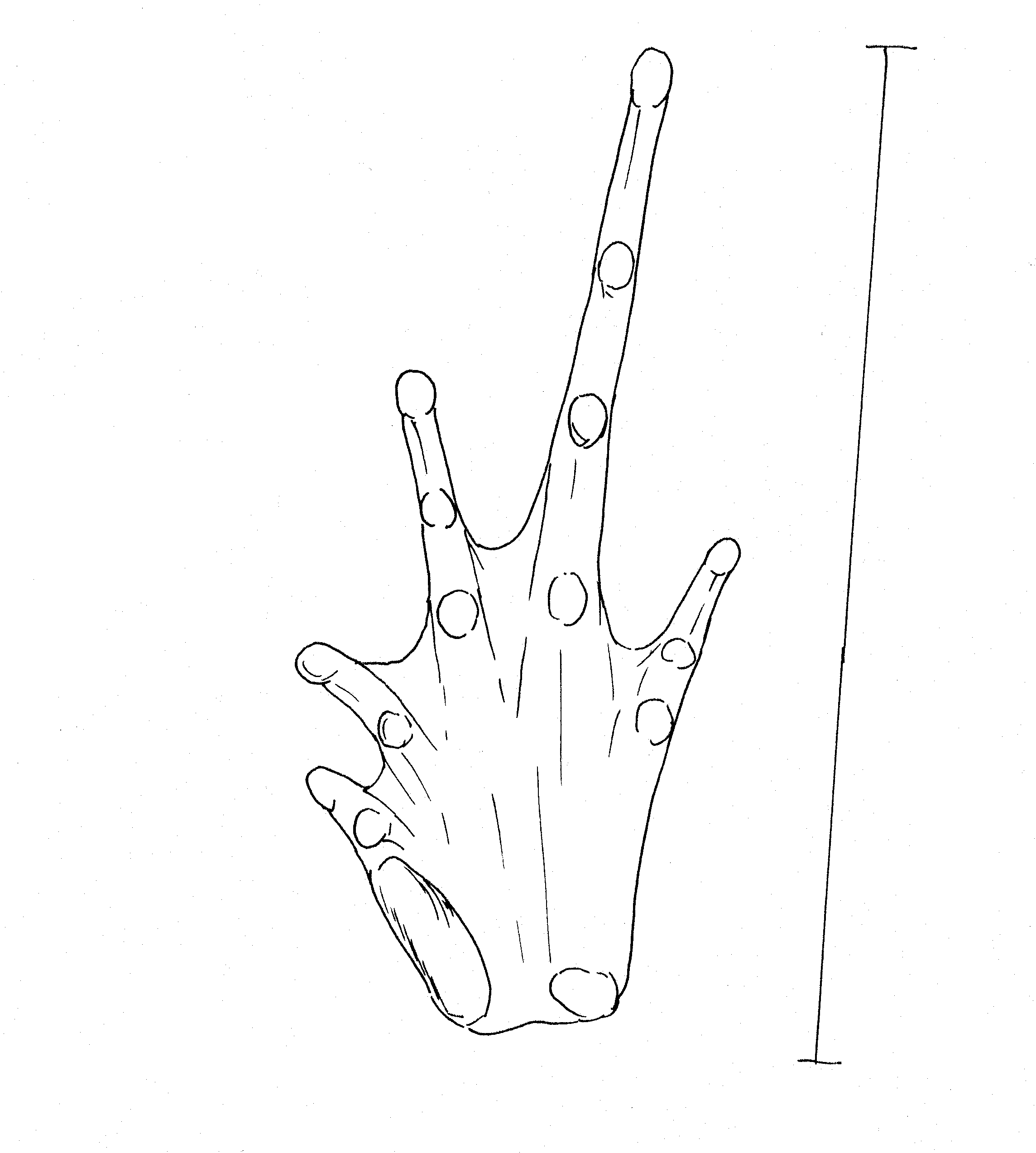
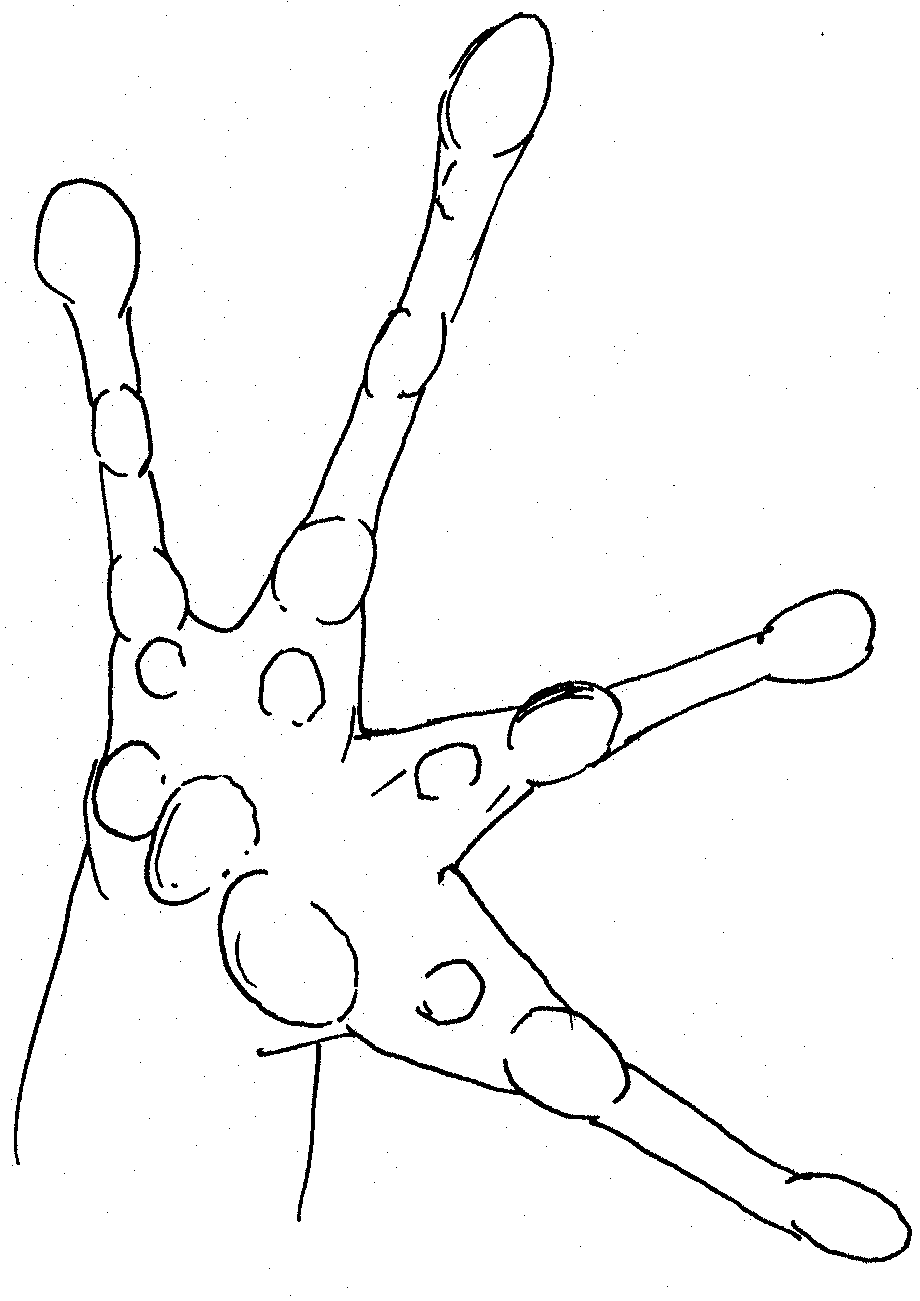
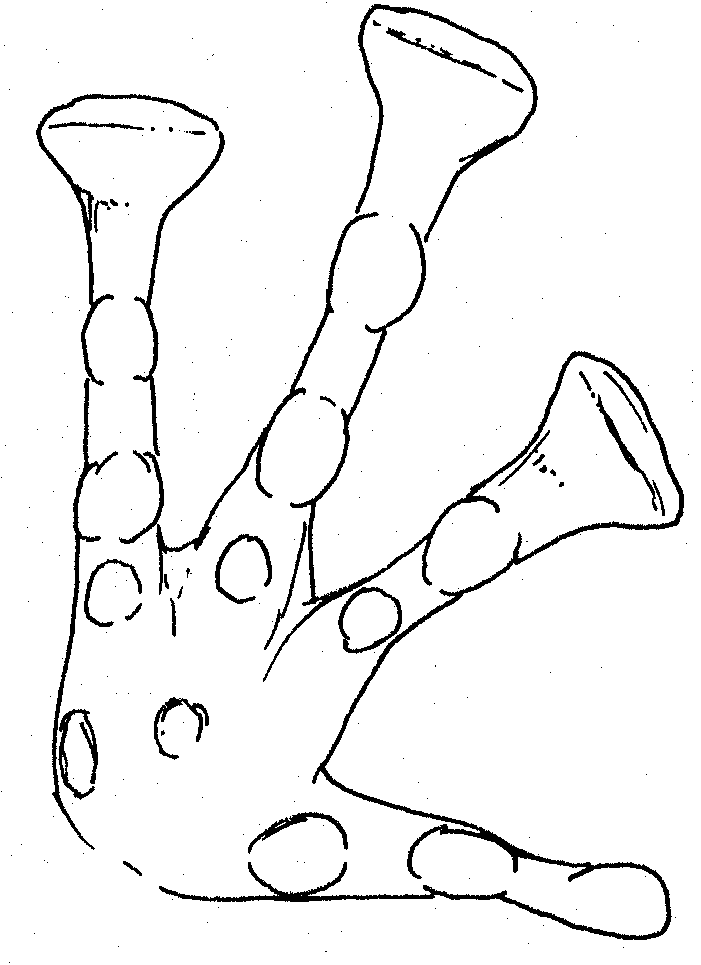 Left
foot of Platymantis sp Right foot of female Platymantis sp.
Large inner metatarsal tubercle, pointed at tip. Very pointed 2nd
tubercle on outer toe
Left
foot of Platymantis sp Right foot of female Platymantis sp.
Large inner metatarsal tubercle, pointed at tip. Very pointed 2nd
tubercle on outer toe
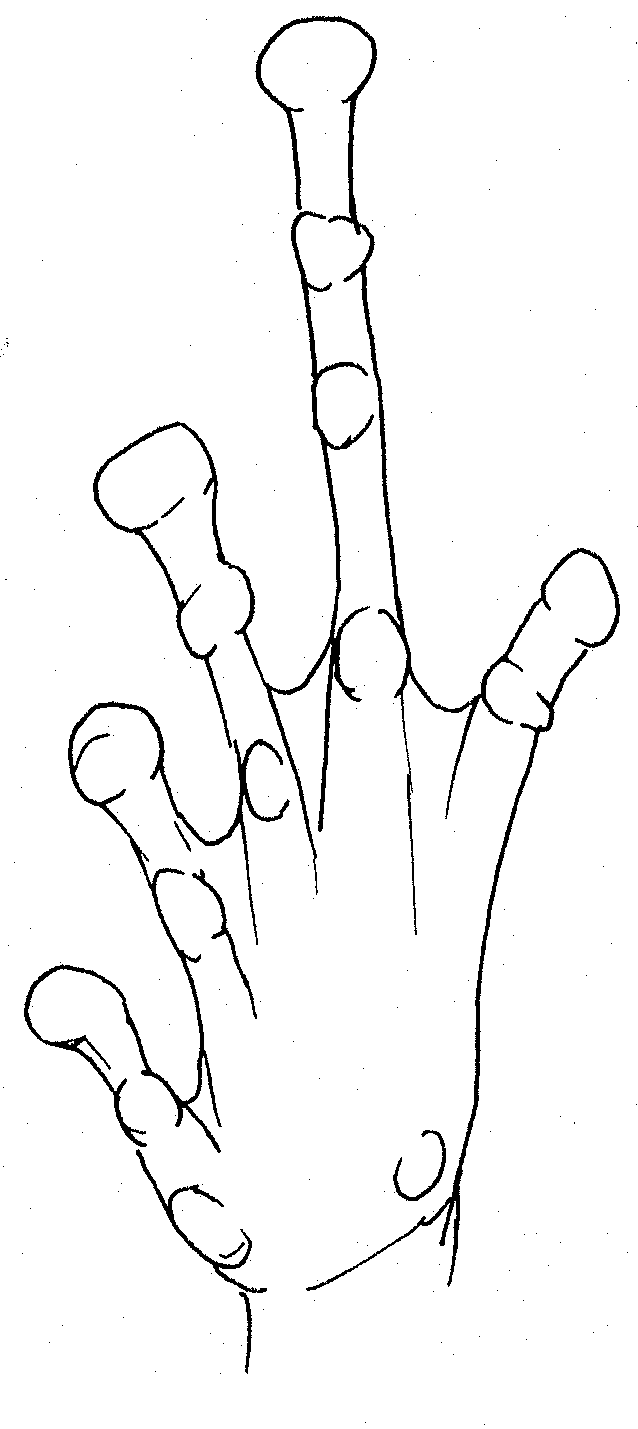
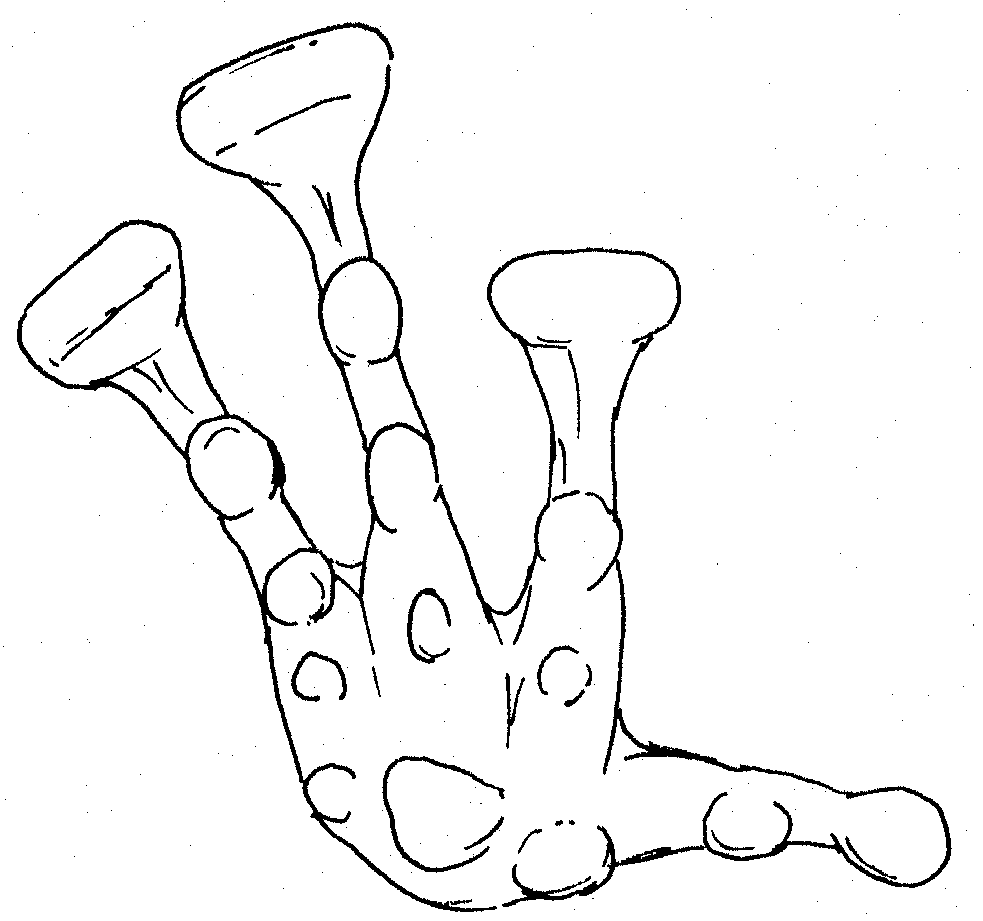 Right hand of male Platymantis
sp.
Right hand of male Platymantis
sp.
![]() Profile of outside finger of right
hand:
Profile of outside finger of right
hand:
1 small tubercle at base of hand, 1 small tubercle at base of finger, 2 large tubercles on finger
The White frog
A small white frog was found on Polillo. It resembles closely Platymantis sierramadrensis, described in 1999 (Brown, W.C., et al. 1999). One individual (0.325g SV 19 mm) was found on 14/8/99, 1m above ground on a leaf next to a stream at site K, Sibulan watershed reserve. On the 9/9/99 a white frog (0.5g SV 22 mm) was found calling (call recorded 2140 hours 25°C) from ferns 2m high above a small stream, North Polillo, near Panukulan. On the 11/9/99 a white frog (0.55g) was heard calling from same location as on the 9/9/99, possibly the same individual.
The table below gives descriptions of the hands and feet of P. polilloensis from Taylor (1922) and Inger (1954), which resemble closely the drawings of the white frog, (see diagram) and comparative descriptions of P. sierramadrensis from (Brown et al. 1999).
|
Inger |
Taylor |
P. sierramadrensis |
|
Tips of fingers broadly dilate into truncate discs with horizontal circummarginal grooves. First finger shorter than second. A supernumerary tubercle on each metacarpal |
Tip of each of the three outer fingers with a strongly dilated pad, nearly twice the width of the digit. Inner finger very small, slender without pad. Subarticular tubercles large, flat, moderately well defined. A single large carpal tubercle. No trace of web |
Fingers with minute webs at base; fingers except first with broad disks; fingers with shallow circummarginal grooves. Finger discs broader than those on toes. First finger shortest, third longest and second and fourth approximately equal, subarticular tubercles large, round, low: a row of low, inconspicuous supernumerary tubercles on palm; inner, middle and outer metacarpal tubercles oval, vague, inner and middle about equal in size; larger than outer. |
|
No supernumerary tubercles on the foot. Tips of toes dilated, the discs smaller than those of fingers; web reaching centre of proximal subarticular tubercle on first, second and fourth toes, distal edge of tubercle on third, and between subarticular tubercles on fifth toe. Web extending to disk of fourth toe as a narrow fringe; third toe slightly longer than fifth; first toe, measured from distal edge of inner metatarsal tubercle, less than twice the length of the tubercle; two metatarsal tubercles, the outer one not distinct; without supernumerary tubercles on the foot. |
Tips of toes distinctly dilated. Not more than one and one half times the width of the digit. Fourth toe very long. Third and fifth equal, barely reaching third subarticular tubercle from the disc on fourth toe. Inner metatarsal tubercle rather large but ill defined. Outer small, dim. |
Terminal phalanges rounded, toes webbed to distal edge of tubercle on first and second, to distal edge of basal tubercle on third, and to midway between tubercles on fifth. Disks of toes narrower than those of fingers, subarticular tubercles rounded, low; platar area smooth; inner metatarsal tubercle elongated, outer vague; dorsum smooth, without tubercles; |
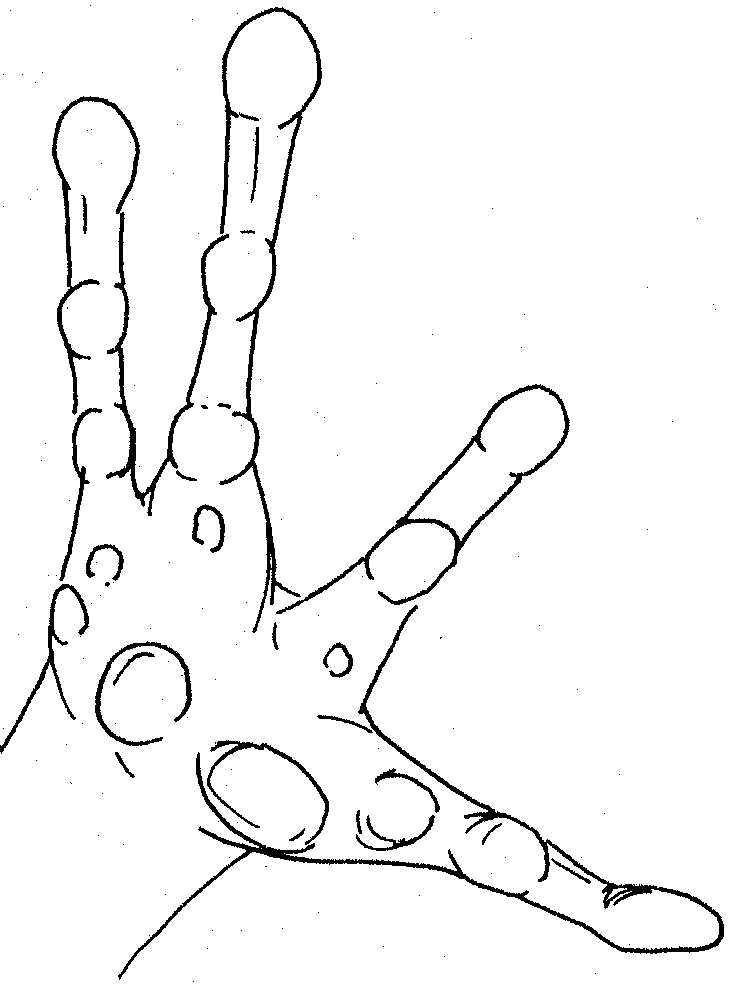
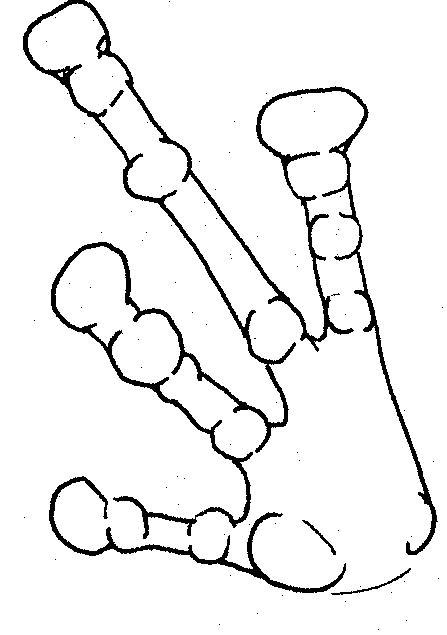
Left hand of white frog showing tubercles and dilated digits
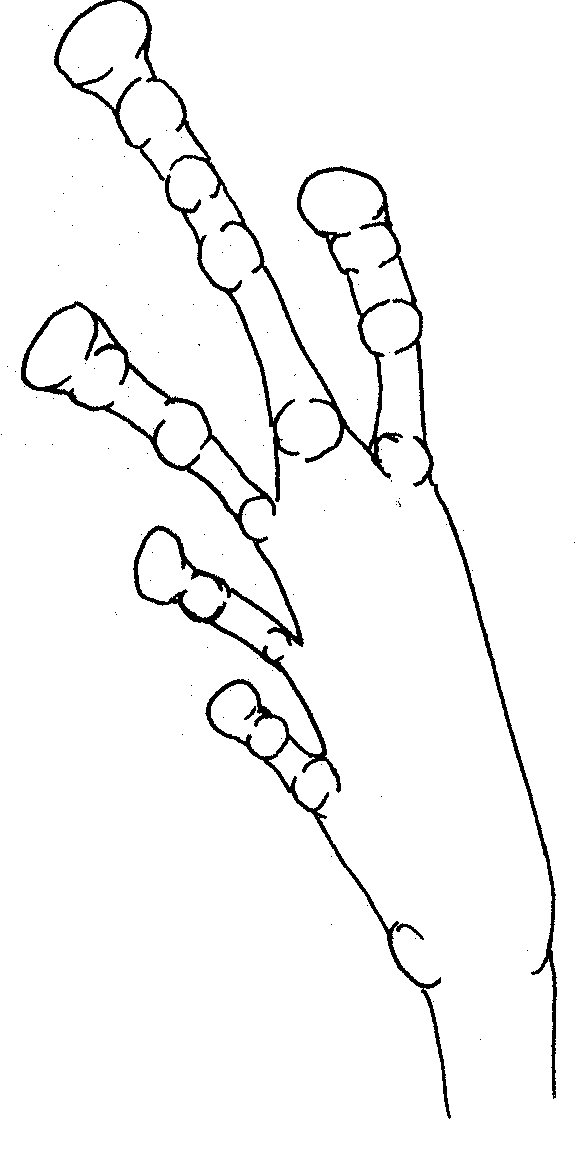 No
apparent webbing Foot of white frog showing tubercles
No
apparent webbing Foot of white frog showing tubercles
|
Description of white frog |
From Taylor 1922: Philautus polilloensis |
P. sierramadrensis from Brown et al 1999. |
|
Prominent tubercle on each eyelid |
Single large conical tubercle on upper eyelid near outer middle edge. Inger: Low papilla on upper eyelid |
|
|
Very pointed tip to snout |
Tip of snout conical. A distinct tubercle on tip of jaw. (Inger: Snout acutely pointed) |
|
|
Pupil horizontal |
Pupil horizontal |
|
|
Small tympanum, one third diameter of eye.
|
Tympanum rather indistinct, diameter 1 third to 1 fourth of eye. (Inger: Tympanum visible, 1 third diameter of eye.) |
|
|
Clear supra tympanic fold, and another fold beneath tympanum, Small fold from tympanum to shoulder |
A strong fold above tympanum to near insertion of arm dimly granular. A second fold from a point above and behind tympanum to lower jaw. There is some variation in the folds about the tympanum, some of these specimens show these very dimly. (Inger: A supratympanic fold from eye to axilla). |
|
|
Prominent tubercle on heel |
A strong tubercle at heel. |
Conical tubercle on heel |
|
Pointed anus |
A small tubercle on back above anus |
|
|
Granular belly |
Belly and underside of femur strongly granular. Skin of throat and belly smooth, with numerous minute pit like depressions. No 349 with Pit like depressions in skin of lower jaw not evident, the tubercles on the back are dimmer. (Inger: Belly with coarse granules) |
Venter generally smooth, coarsely granular in some specimens |
|
See diagram for tubercle positions and pigmentation |
Skin above practically smooth with two small tubercles between shoulders and two pairs on back. (Inger: Skin of back smooth with a few scattered tubercles. No longitudinal ridges). A row of granules from behind tympanum to arm, parallel to supratemporal fold |
smooth skin, dorsum smooth without tubercles. |
|
SV: 22 mm 19 mm |
SV 27 mm female, 22.5, 20, 19, 17, 15. (Inger: male with vocal sacs 20.3 & 22.6 mm SV) |
SV 22.7-25.2; mature males, 25.7 for 1 mature female. |
|
Colour in life: |
Colour in life: Between the 7 co types, vary more or less in markings but the colouration is identical |
|
|
Darker yellow brown under legs, Yellow beneath thighs and hind legs |
Chin and throat yellow with very sparse peppering of brown; underside of leg and foot strongly peppered with brown. (Inger: No areolations on the groin). |
|
|
Thin very pale straight stripe down dorsum |
Numbers 352 to 354 have a hairline from between the eyes to above the anus |
|
|
Above creamy/ off white. Slightly darker pigmented spots as labelled in diagram (dark bar between eyelids and on dorsum), and slightly darker patch on digit. Mask on face from eye to snout |
Above creamy white to yellow, slightly pigmented with minute dots of cinnamon brown. No 350 Almost white when taken. A bar of cinnamon between eyes and dim spots about the dorsal tubercles. No 350 The pigment is heavier between the eyes and as far forward as the line between the eyes. Upper eyelids dark gray to blackish. Dim bars on tibia and femur. |
pale creamy colour without brown/ blackish markings on body or limbs and without areolations. In preserved specimens, faint scattered brownish flecks are evident on dorsal and lateral surfaces |
|
Thin dotted stripe along upper side of each calf, down leg (see diagram), and off white stripe bordering the outside of every finger |
Spots on outer digits of all limbs. The distinct spots on the two outer fingers is very characteristic |
|
|
White belly |
Underside of hand and belly immaculate |
|
|
Iris periphery turquoise |
Schematic drawing to show arrangement of
prominent coloured tubercles.
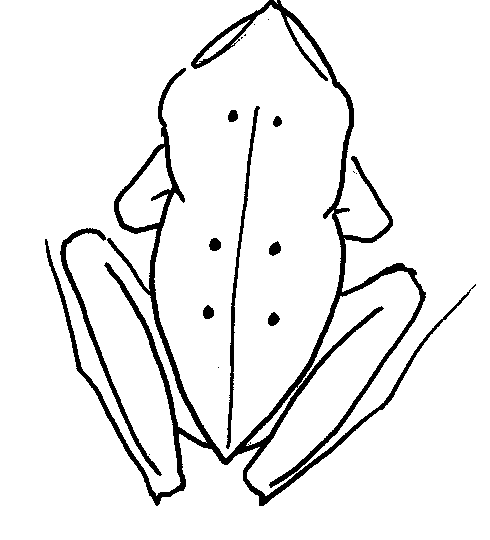
 Right
leg of white frog: Dotted line along back of leg, and fringe of white borders
edge of toe
Right
leg of white frog: Dotted line along back of leg, and fringe of white borders
edge of toe
 Head of Cornufer polilloensis.
(Inger) Head of white frog
Head of Cornufer polilloensis.
(Inger) Head of white frog
Both have pointed snouts, the drawing from Inger does not show tubercle on eyelid but it is given in the description
The description of the white frog corresponds closely to both Inger's and Taylor's descriptions of the original P. polilloensis specimens (referred to as Philautus polilloensis by Taylor and as Cornufer polilloensis by Inger). See accompanying diagrams of the white frog. However the very distinctive turquoise periphery to the iris is not described by Taylor from the original live animals, leaving doubt as to whether the white frog could be P. polilloensis. The blue ring seems to be a character uniting all hazelae group species (R. Brown pers. comm.). The description of P. sierramadrensis also matches the white frog, but their calls are compared in the sound analysis section and found to be distinct. However the comparison is only between calls from populations of P. sierramadrensis in the Northern regions of the Sierra Madre Mountains.
To resolve the identity of the white frog, more specimens are necessary to compare to the original P. polilloensis and P. sierramadrensis specimens, and more recordings of each of the calls are necessary to confirm without doubt calling pattern and rate differences. Further comparison is necessary between the white frog and Southern populations of P. sierramadrensis.
Notes
Eggs presumed to be from a platymantis species were found in a tree hollow. Many gravid platymantis individuals were seen however none could be induced to lay eggs or even call when kept in captivity. Several tiny platymantis were found but neither in great abundance to locate a hatching site nor with any strong association with water. Further investigations into the natural history and reproductive strategies of these species should be investigated.
AN INVESTIGATION INTO THE AMPHIBIAN ASSEMBLAGES OF POLILLO ISLAND, PHILIPPINES
ABSTRACT
A comparison of amphibian assemblages was made between different habitats and management regimes on the island of Polillo, Philippines. A variety of different techniques were used to estimate density and abundance of amphibian species in agriculture, primary and secondary forest and habitat edges. No one technique was found to be suitable for every species, the advantages and disadvantages of each technique are discussed and recommendations for future studies made. A number of species were found to be agricultural specialist species and forest specialist species. Several species were found in all habitats except the most intensive agriculture. Abundance of frogs tends to increase with distance from agriculture. Highest levels of species richness were observed at central forest sites and forest agriculture boundaries where both forest and agriculture species coexist. The forest specialist species were not found at these boundary areas and should be used as a focus of conservation efforts. 16 amphibian species were found and the calls of 12 species were recorded. Specific research and conservation priorities were identified including the taxonomic determination of three unknown species.
INTRODUCTION
Over half the amphibians species on Polillo are endemic to the Philippines. Several are restricted to the South Luzon Faunal region and Platymantis polilloensis, discovered by Taylor in 1922 is found only on Polillo, and has not been found since (Alcala and Brown 1998). Since Taylor’s collecting trips in 1920, only Dr R. Crombie (Smithsonian Institution) has conducted formal herpetofaunal investigations on the island, in the early 1990s. Before the Oxford-UPLB project no acoustic guide to the identification of amphibians on Polillo, nor the Philippines was available, despite recent increases in use of such techniques. This project intended to collect baseline data on amphibian species on Polillo, and produce an aural identification guide.
Amphibians sensitivity to environmental variation suggests they may be affected by habitat and landscape alteration (Wyman 1990; Wake 1991, Blaustein et al.1994). In tropical regions forest fragmentation is an important result of human activity and might influence amphibian assemblages through edge effects on environmental parameters (Ranney 1977; Ranney et al 1981; Lovejoy et al 1986; Malcolm 1994). Most of the forest on Polillo has been cut down, and much of what remains is fragmented. 200 hectares of old growth forest is protected as the Sibulan watershed reserve. Studies of amphibian diversity and richness were conducted in and around the watershed to investigate frog assemblages in different habitats from primary forest to intensive agriculture and begin to address the affect of forest fragmentation on particular species.
AIMS
Methodology
Pilot study:
A week’s reconnaissance and training period was completed at the start of the project. During this time different methods of searching were assessed, different transect lengths and quadrat sizes tested, species identification practised and potential sites investigated. The methodology used in the main study was refined during this period.
Site selection:
Study sites are shown on the map. There was no suitable map of the area available beforehand, thus selection of sites in terms of distance from primary/ core forest was not possible. The arrangement of habitat type and quality was also unknown. The distance to study sites and terrain of routes prevented their random selection. On the assumption that forest nearest to the centre of the reserve (i.e. furthest from agriculture) would be most representative of the original habitat, sites were chosen in what was considered to be those representative of high quality habitat, through varying levels of disturbance to agriculture. The most intensive agriculture was paddy fields.
The project work was based around water bodies because:
Habitat Characterisation:
In order to compare between sites in a quantitative way, different characteristics were measured that were chosen as indicators of forest quality, or factors that may play a role in the distribution of frogs. The following measurements were taken along a distance of 100m, and 5m either side of the river:
To provide a quantitative description of the composition and structure of forest habitats, density, basal area and relative species abundance of trees were estimated at all sites using the Plot less Centre Quarter Method. Distance from origin to the nearest tree (or tree stump) with Circumference at Breast Height >31cm in the four quadrats of a Cartesian grid were recorded together with species, position in the canopy and CBH, every 5m of the transect. Transects were at least 100m long, 50m on each side of the water body and running perpendicular to it. Replicate samples were recorded as such and the distance to next nearest tree measured. Trees were initially identified by local names wherever possible and subsequently by identification of voucher specimens at Dept. of Plant Sciences, University of the Philippines, Los Banos (UPLB). Details of climate (rainfall, maximum and minimum shaded and unshaded temperatures) were recorded from a hill next to base camp at 0900 hours each morning. The phase of the moon during the study period was recorded.
Amphibian Survey Methods:
Two sites were surveyed each night. At each of the 13 study sites on 4 separate occasions the following were conducted:
1. Transects: Twelve 5m by 2m transects were searched. The species, distance from the water, distance from the start of the transect and height above the ground for every frog within the 10m2 was recorded. Six of the transects were directly parallel to the waters edge and six perpendicular to the stream, all 12 beginning at the water’s edge.
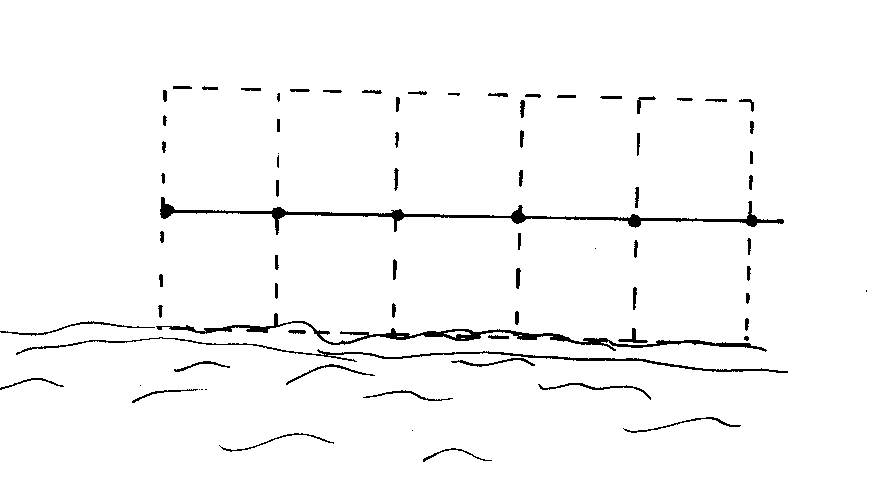
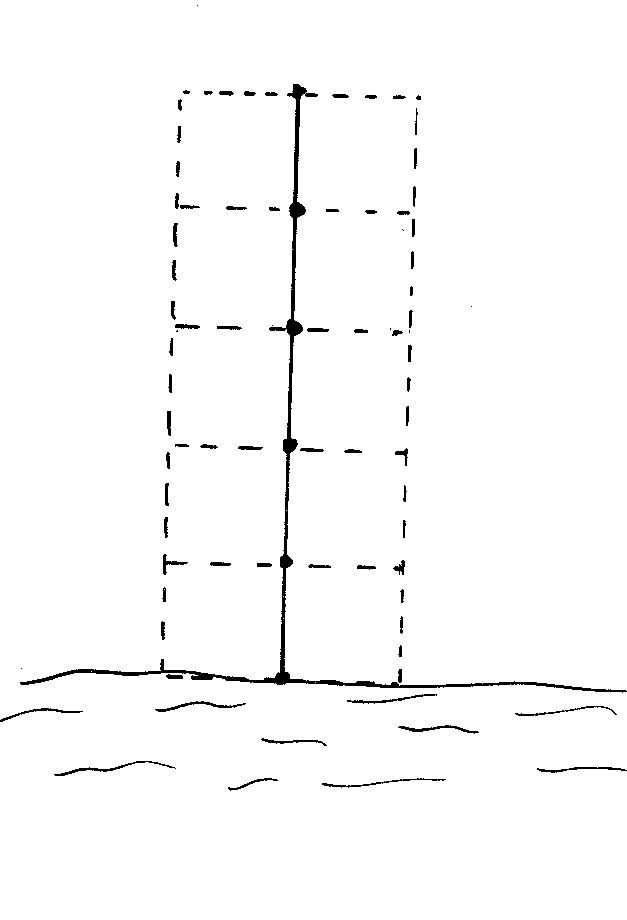 A
5m length of string, marked at 1m intervals was tied to the transect starting
point, to ensure accuracy in measuring distances and consistency between
searches. The rough position of the frog within the 10m2 transect
could be estimated by the 1m markers. Searchers worked from the starting
point keeping the string taut, ensuring no disturbance was caused to the
area ahead. The diagrams below illustrate how knotted string was used to
conduct perpendicular and parallel transects that covered a 10m2
area.
A
5m length of string, marked at 1m intervals was tied to the transect starting
point, to ensure accuracy in measuring distances and consistency between
searches. The rough position of the frog within the 10m2 transect
could be estimated by the 1m markers. Searchers worked from the starting
point keeping the string taut, ensuring no disturbance was caused to the
area ahead. The diagrams below illustrate how knotted string was used to
conduct perpendicular and parallel transects that covered a 10m2
area.
2 researchers searched transects simultaneously at each site. Each searched 6 transects; 3 parallel and 3 perpendicular. The searchers worked independently and on opposite sides of the water body to avoid pseudoreplication. Transects were at least 5m apart to prevent active frogs being included in consecutive transects.
Searching began between 6.50 - 7.30pm. Searching was thorough and slow, the time taken to complete a transect was left to the searchers’ discretion, given the varying terrains and abundances of frogs. Searchers worked at a pace, where they felt, given more time, unable to find any more frogs within the search area. If a frog was seen to jump from outside the transect area into the area it was not counted, if a frog was seen to jump out of the transect area it was counted. Times to complete a transect varied between 2 to 20 minutes.
The transects were designed to give absolute abundances of every species, to enable patterns in species distributions with distance from the waters edge to be deciphered and to look for signs of clustered distributions.
2. Timed counts: For 30 minutes searchers looked anywhere in the vicinity of the area. No constraints were given to search area dimensions other than that searchers worked on opposite sides of the river from each other to avoid replication. The species of the frogs found were tallied, and the time noted when 10 individuals of 1 species had been found. With the aim of saving time that would be used up by continuing to record the common species (perhaps increasing the possibility of finding more cryptic species). Timed counts are designed to be a simple, rapid and effective technique which gives reasonable relative abundances for avifauna (Bibby 1998). Therefore an adapted version of the technique was tested for assessing frog assemblages.
Ten different searchers carried out the surveys. They were chosen on a rota basis to reduce observer bias, random selection was impossible because of the logistics of the work timetable, frequency of fungused feet and other ailments preventing work. Petzl head torches were used by searchers. Searches were not completed on Sundays and were abandoned when dangerous weather conditions were anticipated. Therefore a total of 48 transects and four hours of timed counts were completed at each site. The advantages and disadvantages of these two techniques are reviewed in the discussion.
Analysis methods:
Species accumulation curves were drawn to assess what proportion of the species present were being sampled. Rank abundance plots were used to assess overall species community patterns and identify habitat specialists. Graphs of species richness and total frog abundance for each site are presented and diversity indices calculated. Ordination using the programme CAP was used to look for patterns in species composition and abundance between sites. Density of each frog species is plotted against distance from agriculture.
Considerations in Design of Methodology:
Of the recommended standard techniques for sampling amphibian populations (Heyer et al 1994) few were considered appropriate for the community being studied on Polillo. Quadrats typically 5m by 5m were rejected because they:
Long transects (100m plus) were rejected because
However a compromise upon both the quadrat and transect design was reached. Small 5m long, 2m wide transects were used that could be searched intensively and repeated many times. Surveying was purely conducted at water bodies, but a small gradient (5m) was used perpendicular from the water bodies to investigate whether on future occasions further work based on the forest floor would be worthwhile.
Considerations about the design of the transects:
Differences in searcher ability are very difficult to overcome and impossible to eliminate. Differences were minimised by the design of the experiment wherever possible. E.g. by keeping the search area small (5m by 2m), by working at the searchers slowest pace (to ensure searchers didn't feel rushed thereby likely to search less thoroughly), by working in one direction (thereby any frogs that jump in front are likely to be seen). A larger search area was considered to increase searcher boredom and apathy, less successful searchers might become disheartened, heightening differences between more successful searchers. (It was assumed that given enough time the searcher will find all the visible frogs no matter what ability the searcher has, given a small enough area). The area searched was small to ensure that if frogs jumped they were likely to be seen. The design was a transect rather than a quadrat to reduce the searcher disturbance and likelihood of frogs escaping before they were spotted. 2m width was considered to be a good field of vision for the observers.
Considerations about the design of the timed counts:
This method tended to be more popular during the reconnaissance period, therefore it was important to consider whether this gave the same relative levels of each species as the transect method (although area is not specified therefore neither relative or absolute abundances can be measured, only frequency of encounters). It was assumed that searchers would try to find as many frogs as possible. A limit to the number of frogs of one species recorded was made, to prevent the tendency to concentrate efforts on one species or productive microhabitat. After trying different limits, 10 frogs of one species was chosen. In order for appropriate analysis, the number of frogs that would have been found in thirty minutes was extrapolated given the encounter rate for ten frogs.
The two search techniques were suspected to collect differing results. E.g. The timed count might not pick up species that are particularly cryptic since this method is possibly less intensive. However the search area and habitats are possibly wider therefore there is more chance of finding animals with lower densities.
Audio strip transects were latterly adopted with appropriate changes for estimating density of Platymantis sp. This technique is not appropriate for many of the other species such as P. dorsalis which have a high call overlap (high densities of males calling at high rates) (Zimmerman 1994) or others such as O. laevis which call in choruses. For Platymantis sp. the technique was ideal. However as densities increase, the technique requires more concentration and skill and is more time intensive therefore shorter and shorter transects must be completed.
Description of Study sites:
D- Central forest site with a medium sized stream (1 to 2m) with medium flow rate.
J- Forest site with small stream. Varying terrain (steep sided in places, flat in others) which affects flow rate (varies from fast to slow).
The following map of the watershed and surrounding area shows the position of the frog surveys sites listed above.

RESULTS
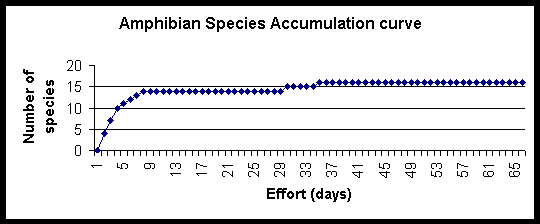
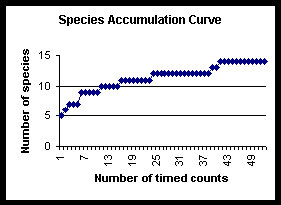
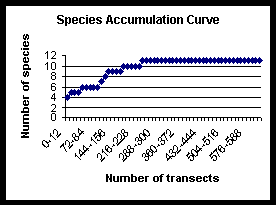
The species accumulation curves compare species richness and rate of species discovery for transect and timed count techniques. The total species richness approaches an asymptote at 16, species were still being discovered after 37 days. The transect technique found 11 of the total 16 species, whereas the timed counts found 14. The two species not found by either technique were seen once and twice respectively (see below and species accounts section). The transect data reached an asymptote one third of the way through the study, whereas the timed count curve continued to rise, although slowly at the end.
Two frog species were only not detected during the surveys; Kaloula.sp, a fossorial frog (2 individuals were found, 1 in a pitfall trap after rain, and 1 during the reconnaisance period approximately 5m from the stream) and Rhacophorus appendiculatus (found on only one occasion at a site within the watershed reserve). Based on call recognition, the species found regularly at significant distances from water were particularly cryptic, and could often only be located by their call, these were Platymantis sp, P. corrugatus, P.dorsalis. Individuals of other species were found at a distance from water, such as R.woodworthi, but only occasionally compared to their abundance at stream sites.
Species richness plotted in rank order for the two techniques does not follow a clear pattern. The forest sites tend to have higher species richness than the paddyfield sites.
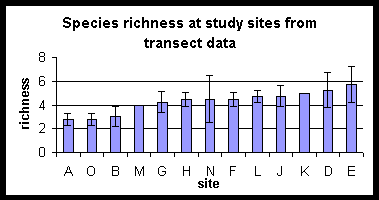
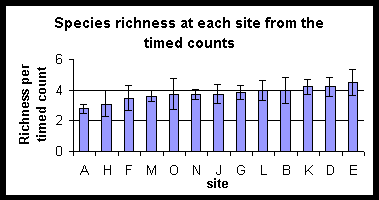 Sites
are ranked in order of increasing species richness. Differences in the rank
order produced by the two techniques should be noted.
Sites
are ranked in order of increasing species richness. Differences in the rank
order produced by the two techniques should be noted.
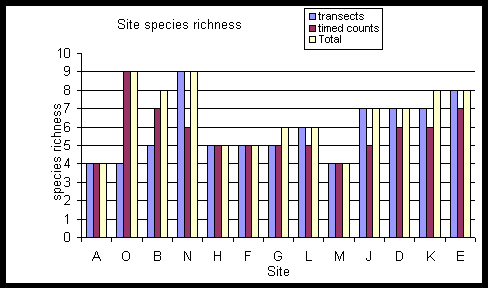 There is no clear distinction
between forest and forest edge sites, shown clearly by the diagram combining
richness estimates from both techniques with sites ranked from agriculture
(A) to the most central forest areas (E). No inference is made about species
known to be present but not detected by the sampling techniques.
There is no clear distinction
between forest and forest edge sites, shown clearly by the diagram combining
richness estimates from both techniques with sites ranked from agriculture
(A) to the most central forest areas (E). No inference is made about species
known to be present but not detected by the sampling techniques.
Sites are ranked in order of their proximity to agriculture. A is a paddy field site and E is the most central forest site.
Rank Abundance Plots using both the transect and timed count results in turn have been produced for all the sites (all ranked based upon the rank abundance for site E, (i.e. the site nearest the centre of the watershed reserve). The pattern in species composition and relative abundance between the two methods is very similar. No meaningful measures of abundance can be calculated for the three species found in timed counts but not in transects, although they must be rare, cryptic or clustered in distribution. Further explanation as to their ecology is given in the species accounts section. All the forest sites have similar rank abundance plots. Both B and N which border agriculture show a larger species richness than both the paddy field sites and some of the forest sites. The diagrams on the following page summarise the Rank Abundance plots for the arbitrararily chosen habitat types; Paddy fields (sites A and O), coconut plantation (sites N and B), forest edge sites (sites H, F, G, L) and central forest (sites M, K, J, D, E).
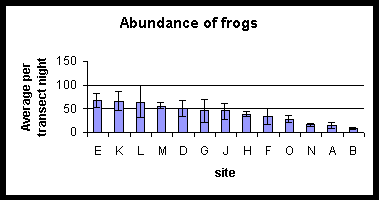
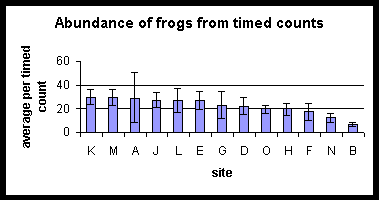
Sites are ranked in order of decreasing abundance of frogs. Changes in order for the two different techniques should be noted.
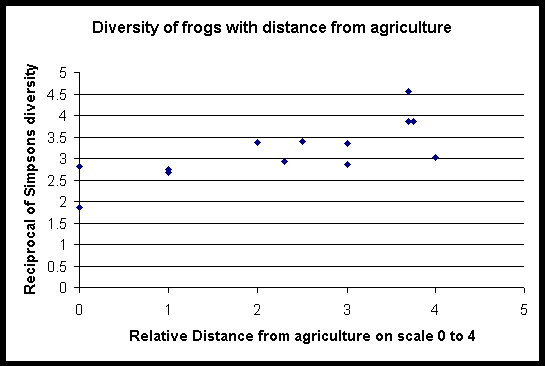
A variety of different diversity indices (Simpson’s index, The Shannon Weiner Function, Brillouin’s Index of species diversity and Evenness measures) for each site were calculated using the transect results. All indices produced similar patterns of diversity for the sites. Central forest sites were the most diverse, and paddy field sites the least. Sites that bordered agriculture or forest edge were more diverse than both the paddyfield sites and several forest sites.
Abundance of all the common species of frog (Rana woodworthi, Rana signata similis, Occidozyga laevis, Platymantis dorsalis) increased with distance into forest. Central forest sites have the highest abundances of frogs. Site L is an exception, but it does have characteristics that make it a good frog habitat e.g. still but permanent water and lots of pandanus plants. Bufo marinus, Rana vittigerra, and Polypedates leucomystax were found exclusively in agriculture. Very few Platymantis sp, Platymantis corrugatus, Rana magna and Rana everetti were found by either technique.
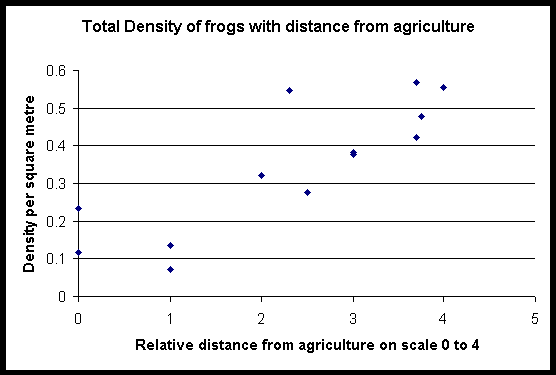
Abbreviations for the amphibian species in all the diagrams are as follows:
Bufo marinus Bm
Rana vittigerra Rv
Rana woodworthi Rw
Rana similis Rs
Polypedates leucomystax Pl
Platymantis dorsalis Pd
Platymantis sp. Psp
Platymantis corrugatus Pc
Limnonectes macrocephalus Lm
Occidozyga laevis Ol
Rana luzonensis Rl
White frog Wf
Rhacophorus appendiculatus Ra
Rhacophorus pardalis Rp
Kaloula picta Kp
Kaloula sp Ksp